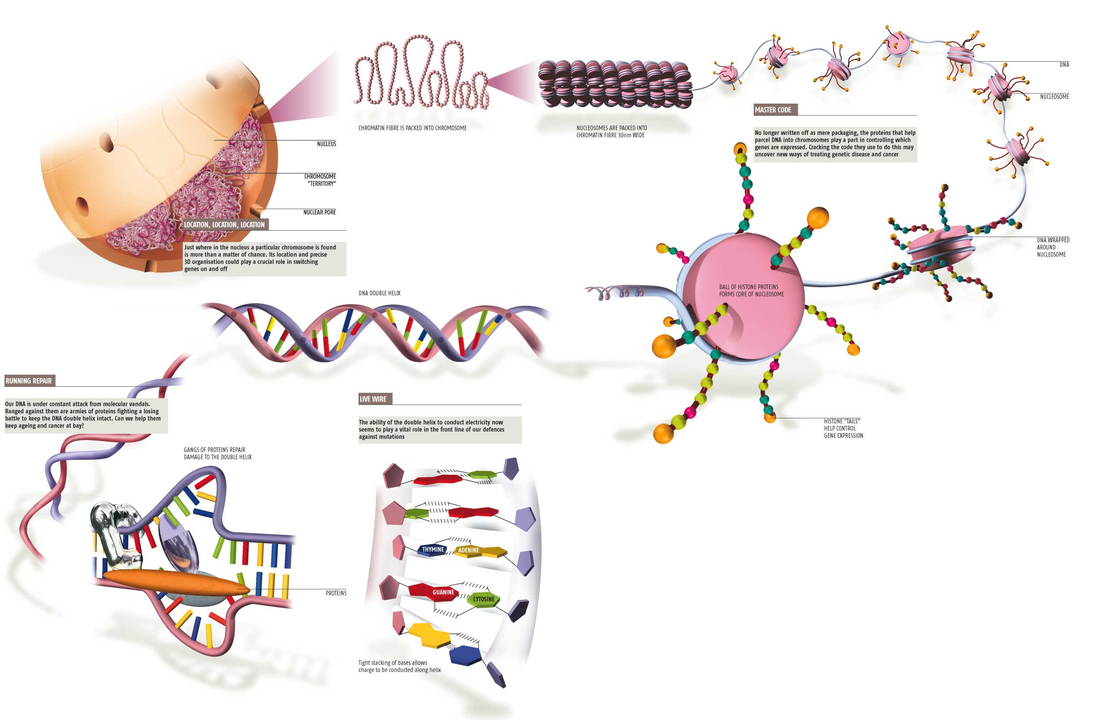
Structural Organization of the Genome
The behaviour of a cell may be likened to a musical rhythm which repeats in a cyclic manner, yet can respond to ques from the immediate environment. The forces which regulate eukaryotic gene expression are both cell autonomous, and non-autonomous, and the consequences of these forces are the behaviours which we seek to understand and ultimately control as molecular biologists. To understand this rhythm which manifests as the regulation of gene expression, one must first understand how the genome is organized.
Chromatin: Structural Organization
In the nucleus of eukaryotic cells, the entire genome is wrapped around histone proteins which compact the DNA into a smaller volume. In mammalian cells, this structural organization allows approximately 2 m of linear DNA to be packed into a nucleus of roughly 10 µm diameter. This combination of DNA and proteins within the nucleus of a cell is referred to as chromatin. In general terms, there are three levels of chromatin organization:
The basic repeat element of chromatin is the nucleosome, interconnected by sections of "linker DNA". A nucleosome is the basic unit of DNA packaging in eukaryotes, consisting of a segment of DNA wound in sequence around eight core histone proteins. Nucleosomes are folded through a series of successively higher order structures to eventually form a chromosome; this both compacts DNA and creates an added layer of regulatory control, which ensures correct gene expression.
The nucleosome core particle consists of approximately 147 base pairs of DNA wrapped in 1.67 left-handed superhelical turns around a histone octamer consisting of 2 copies each of the core histones H2A, H2B, H3, and H4. All four of the histones that make up the core of the nucleosome are relatively small proteins (102–135 amino acids), and they share a structural motif, known as the histone fold, formed from three α helices connected by two loops. In assembling a nucleosome, the histone folds first bind to each other to form H3–H4 and H2A-H2B dimers, and the H3–H4 dimers combine to form tetramers. An H3–H4 tetramer then further combines with two H2A-H2B dimers to form the compact octamer core, around which the DNA binds.
Adjacent nucleosomes are joined by a stretch of free DNA termed "linker DNA" which varies from 10 - 80 bp in length depending on species and tissue type. Technically, a nucleosome is defined as the core particle plus one of these linker regions; however the word is often synonymous with the core particle.In addition to the core histones, there are "linker histones", such as H1 and its isoforms, which sit at the base of the nucleosome near the DNA entry and exit, and associate with "linker DNA". Linker histones are involved in chromatin compaction. Under non-physiological conditions, non-condensed nucleosomes with about 20 to 60 base pairs of linker DNA can form without the linker histone, and form a 10 nm "beads on a string" fibre of DNA.
Dynamic fluctuations in chromatin structure allow for the expression or repression of genes throughout the genome. Generally, the "relaxed" structure of euchromatin is transcriptionally permissive due to unobstructed access to gene promoters for the transcriptional machinery. In contrast, the "compact" structure of heterochromatin (histones wrapped into a 30 nm fiber) is transcriptionally repressive due to inaccessibility of gene promoters for the transcriptional machinery. The processes that regulate transcription through modulation of chromatin structure are discussed in subsequent sections.
Anjana Rao 2019:
Principal component analysis of the interaction matrix obtained from Hi-C data has been used to compartmentalize the genome into an A compartment (positive PC1 values) and a B compartment (negative PC1 values) that exhibit the hallmark characteristics of euchromatin and heterochromatin, respectively. The euchromatic A compartment is rich in expressed genes in the cell type under consideration, whereas the heterochromatic B compartment is gene-poor and bears epigenetically “repressive” chromatin marks, including H3K9me2/3 (17, 18). Moreover, the Hi-C B compartment overlaps with lamina-associated domains and corresponds to late-replicating regions of the genome, whereas the Hi-C A compartment corresponds to early replicating genomic regions and is not lamina associated (18, 19).
Chromatin: Structural Organization
In the nucleus of eukaryotic cells, the entire genome is wrapped around histone proteins which compact the DNA into a smaller volume. In mammalian cells, this structural organization allows approximately 2 m of linear DNA to be packed into a nucleus of roughly 10 µm diameter. This combination of DNA and proteins within the nucleus of a cell is referred to as chromatin. In general terms, there are three levels of chromatin organization:
- DNA wraps around histone proteins forming nucleosomes; the "beads on a string" structure (euchromatin).
- Multiple histones wrap into a 30 nm fibre consisting of nucleosome arrays in their most compact form (heterochromatin).
- Higher-level DNA packaging of the 30 nm fibre into the metaphase chromosome (during mitosis and meiosis).
The basic repeat element of chromatin is the nucleosome, interconnected by sections of "linker DNA". A nucleosome is the basic unit of DNA packaging in eukaryotes, consisting of a segment of DNA wound in sequence around eight core histone proteins. Nucleosomes are folded through a series of successively higher order structures to eventually form a chromosome; this both compacts DNA and creates an added layer of regulatory control, which ensures correct gene expression.
The nucleosome core particle consists of approximately 147 base pairs of DNA wrapped in 1.67 left-handed superhelical turns around a histone octamer consisting of 2 copies each of the core histones H2A, H2B, H3, and H4. All four of the histones that make up the core of the nucleosome are relatively small proteins (102–135 amino acids), and they share a structural motif, known as the histone fold, formed from three α helices connected by two loops. In assembling a nucleosome, the histone folds first bind to each other to form H3–H4 and H2A-H2B dimers, and the H3–H4 dimers combine to form tetramers. An H3–H4 tetramer then further combines with two H2A-H2B dimers to form the compact octamer core, around which the DNA binds.
Adjacent nucleosomes are joined by a stretch of free DNA termed "linker DNA" which varies from 10 - 80 bp in length depending on species and tissue type. Technically, a nucleosome is defined as the core particle plus one of these linker regions; however the word is often synonymous with the core particle.In addition to the core histones, there are "linker histones", such as H1 and its isoforms, which sit at the base of the nucleosome near the DNA entry and exit, and associate with "linker DNA". Linker histones are involved in chromatin compaction. Under non-physiological conditions, non-condensed nucleosomes with about 20 to 60 base pairs of linker DNA can form without the linker histone, and form a 10 nm "beads on a string" fibre of DNA.
Dynamic fluctuations in chromatin structure allow for the expression or repression of genes throughout the genome. Generally, the "relaxed" structure of euchromatin is transcriptionally permissive due to unobstructed access to gene promoters for the transcriptional machinery. In contrast, the "compact" structure of heterochromatin (histones wrapped into a 30 nm fiber) is transcriptionally repressive due to inaccessibility of gene promoters for the transcriptional machinery. The processes that regulate transcription through modulation of chromatin structure are discussed in subsequent sections.
Anjana Rao 2019:
Principal component analysis of the interaction matrix obtained from Hi-C data has been used to compartmentalize the genome into an A compartment (positive PC1 values) and a B compartment (negative PC1 values) that exhibit the hallmark characteristics of euchromatin and heterochromatin, respectively. The euchromatic A compartment is rich in expressed genes in the cell type under consideration, whereas the heterochromatic B compartment is gene-poor and bears epigenetically “repressive” chromatin marks, including H3K9me2/3 (17, 18). Moreover, the Hi-C B compartment overlaps with lamina-associated domains and corresponds to late-replicating regions of the genome, whereas the Hi-C A compartment corresponds to early replicating genomic regions and is not lamina associated (18, 19).